 Metadata catalogue
Metadata catalogue
Biological oceanography
Type of resources
Available actions
Topics
INSPIRE themes
Keywords
Contact for the resource
Provided by
Years
Formats
Representation types
Update frequencies
-
Sea fans (order : Alcyonacea) are the most remarkable and easily identifiable species of the subtidal hard substrate communities (Gili and Ros, 1985). From an ecological point of view, sea fans play an essential role (Gili and Coma, 1998) by increasing the biomass and the diversity of hard substrates (Mitchell et al, 1992; Ballesteros, 2006) via an umbrella species role providing habitat for small epifauna and a refuge for many fish (Ross and Quatrini, 2007). When their density is high enough, they form animal forests (sensu Rossi et al 2017) and become engineering species by modifying the level of turbulence and therefore of sedimentation of propagules in the benthic boundary layer (Chamberlain and Graus, 1975). In the shallow rocky habitats of the Mediterranean, five species of sea fans dominate: the white gorgonian Eunicella singularis (Esper, 1791), the orange gorgonian Leptogorgia sarmentosa (Esper, 1789), the yellow gorgonian Eunicella cavolinii (Koch, 1887), the red gorgonian Paramuricea clavata (Risso, 1826), and the red coral Corallium rubrum (Linnaeus, 1758). Some of these species, characterized by long life span, have been included in the management plans of the Gulf of Lion Marine Protected Areas, with surveys for the monitoring of the demographic structure of E. Singularis (Réserve Naturelle Marine de Cerbère-Banyuls, Aire Marine Protégée Agathoise) and C. rubrum (Réserve Naturelle Marine de Cerbère-Banyuls, Parc Marin de la Côte Bleue). In parallel with these management-related surveys, some scientific studies on the demographic structure and ecology of P. clavata, E. singularis and C. rubrum, with the aim of documenting mass mortality events (Cerrano et al. al., 2000, Garrabou et al., 2001) have been undertaken in the Parc Marin de la Côte Bleue, in the Parc National de Port-Cros, along the Côte Vermeille and Cap de Creus coast and in the Cinque Terre National Park (Garrabou and Harmelin, 2002, Torrents et al., 2005 ; Linares et al. 2008 Linares et al., 2010 ; Santangelo et al., 2011; Gori et al., 2011a ; Gori et al. 2011b; Rossi et al., 2008). The present database gathers two extensive inventory of sea fans populations performed between 2013 and 2015 in the Gulf of Lion and between 2018 and 2019 in the Ligurian Sea. The same protocol was applied to estimate the population density of the five species in 585 stations. The 585 stations were defined a priori on a regular mesh mapping the main hard-bottom substrate units of the Gulf of Lion (Côte Bleue, Plateau des Aresquiers, Plateau du Cap d'Agde, Cap Leucate, Côte Vermeille), and surrounding the ports of Toulon, La Spezia and Bastia (Figure 1). The spacing between stations was varied from 100m to 800 m according to bathymetry steepness. Each station was geo-referenced from the surface and located on the sea bed with a mooring. Counts of individuals of the five species were made by scuba divers trained to species identification in four quadrats (1m x 1m), positionned at 5 m from the mooring along the sea bed, in the four cardinal directions. In total, the sampling required 1500 dives. References Ballesteros E. 2006. Mediterranean coralligenous assemblages: a synthesis of present knowledge Oceanography and Marine Biology: An Annual Review 44, 123-195 Carpine C, Grasshoff M. 1975. Les gorgonaires de la Mediterranee. Bull Inst Oceanogr Monaco 71:1–140 Cerrano, C., Bavestrello G., Bianchi C.N., Cattaneo R. Vietti, S. Bava, C. Morganti, C. Morri, P. Picco, G. Sara S. Schiaparelli S. Siccardi A. & Sponga, F. 2000. A catastrophic mass-mortality episode of gorgonians and other organisms in the Ligurian Sea (Northwestern Mediterranean), summer 1999. Ecology Letters 3: 284–293. Chamberlain J.J. A., Graus, R.R. 1975. Water Flow and Hydromechanical Adaptations of Branched Reef Corals. : Bulletin of Marine Science 25 (1): 112-125 Garrabou J., Perez T., Sartoretto S., Harmelin J.G., 2001. Mass mortality event in red coral Corallium rubrum populations in the Provence region (France, NW Mediterranean). Mar Ecol Prog Ser 217:263–272 Garrabou J., Harmelin G. 2002. A 20-year study on life-history traits of a harvested long-lived temperate coral in the NW Mediterranean: insights into conservation and management needs. J Anim Ecol 71:966–978 Gili, J.M., & Ros, J. 1985. Study and cartography of the benthic communities of the Medes Islands (NE Spain). Marine Ecology 6, 219–238. Gili J.M. and Coma R. 1998. Benthic suspension feeders: their paramount role in littoral marine substrates. TREE 13 (8): 316-321 Gori, A., Rossi, S., Linares, C., Berganzo, E., Orejas, C., Dale, M. & Gili, J.M. 2011a. Size and spatial structure in deep versus shallow populations of the Mediterranean gorgonian Eunicella singularis (Cap de Creus, Northwestern Mediterranean Sea)” Marine Biology, DOI: 10.1007/s00227-011-1686-7 Gori A., Rossi S., Berganzo E., Pretus J.L., Dale M.R.T., Gili J.M. 2011b. Spatial distribution patterns of the gorgonians Eunicella singularis, Paramuricea clavata and Leptogorgia sarmentosa (Cape of Creus, Northwestern Mediterranean Sea). Mar Biol 158:143-158 Linares C, Coma R, Garrabou J, Diaz D, Zabala M. 2008. Size distribution, density and disturbance in two Mediterranean gorgonians: Paramuricea clavata and Eunicella singularis. J Appl Ecol 45:688–699 Linares C., Bianchimani O., Torrents O., Marschal C., Drap P., Garrabou J., 2010. Marine Protected Areas and the conservation of long-lived marine invertebrates : the Mediterranean red coral. Marine Ecology Progress Series 402: 69-79. Mitchell, N. D., Dardeau, M. R., Schroeder,W. W., Benke, A. C. 1992. Secondary production of gorgonian corals in the northern Gulf of Mexico. Mar Ecol. Prog. Ser. 87: 275-281 Ross S., Quattrini A. 2007. The fish fauna associated with deep coral banks off the southeastern United States. Deep Sea Research I (54): 975-1007. Rossi S, Bramanti L, Gori A, Orejas C. 2017. An Overview of the Animal Forests of the World. In book: Marine Animal Forests, pp.1-26. SPRINGER.[DOI: 10.1007/978-3-319-17001-5_1-1] Santangelo G., Bramanti L., Rossi S., Tsounis G., Vielmini I., Lott C., Gili J.M. 2011. Spatial patterns of variation in recruitment and post-recruitment processes of the Mediterranean precious gorgonian coral Corallium rubrum. Journal of Experimental Marine Biology and Ecology. DOI: 10.1016/j.jembe.2011.10.030 Torrents O., Garrabou J., Marshal C., Harmelin J.G., 2005. Age and size at first reproduction in the commercially exploited red coral Corallium rubrum (L.) in the Marseilles area (France, NW Mediterranean). Biol Conserv 121: 391–397 Rossi S., Tsounis G., Orejas C., Padrón T., Gili J.M., Bramanti L., Teixidó N. and Gutt J. 2008. Survey of deep-dwelling red coral (Corallium rubrum ) populations at Cap de Creus (NW Mediterranean). Marine Biology 154 (3): 533-545.
-
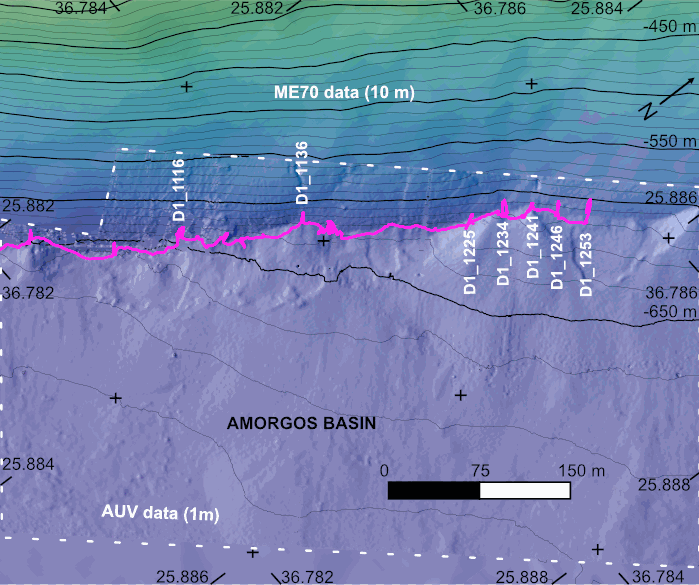
The AMORGOS-23 cruise (https://doi.org/10.17600/18003211) was conducted onboard R/V Europe from April 26th 2023 to May 9th 2023. Using HROV Ariane equipped with 4K cameras, we surveyed the main active faults between Amorgos and Anafi islands in order to identify and quantify recent deformation of the seafloor linked to the 1956 Amorgos earthquake. During dive 244-1, we surveyed the steepest scarp of the Amorgos fault, along its central segment. We provide here seven scaled Digital Outcrop Models (in .obj format) calculated from the HROV videos with MATISSE 3D software. There location (model origin) is also provided in a table, together with information about the portions of the videos used to produce the models. Due to incomplete navigation data, the models are not properly oriented.
-
Data of parameters presented as figures in the manuscript: Biogenic silica (BSi: µmol.L-1), Lithogenic silicon (LSi: µmol.L-1), Total Chlorophyll a (TChla: mg.m-3) and Fucoxanthin (Fuco: mg.m-3). Note that: - Total fraction for BSi and LSi is available from Niskin bottles (> 0.8 µm) and in situ pumps (> 0.45 µm); - Size-fractions for BSi and LSi is available from in situ pumps only (5-53 µm; > 53 µm); - Total fraction for TChla and Fuco is available from Niskin bottles (> 0.7 µm). - Integrated data (0-200 m) is available from Niskin Bottles. Acronym explanations: - Sampling type: ISP = In Situ Pump; ISP-SF = In Situ Pump with Size-Fraction; NSK = Niskin. - Method used for BSi and LSi computation: NOC = No Correction applied; RAG = Correction using the method used from the protocol established by Ragueneau et al. (2005); ACR = Average Crustal Ratio method used when one of the criterion from the protocol established by Ragueneau et al. (2005) is not respected. - bdl = Below Detection Limit For further explanations in the method, should you please refer to the Material and Methods section in the manuscript (revised version submitted in Marine Chemistry).
-
Black and white seafloor photomosaic of the Lucky Strike hydrothermal field and the ridge axis to the South, acquired with VICTOR6000 during the Bathyluck 2009. Individual still images were matched, corrected for uneven illumination, renavigated and blended for a seamless photomosaic. This data release includes partial geotiffs covering the whole surveyed area. To be cited: Gini et al. (submitted); Barreyre et al. (2012); Escartin et al. (2013); Escartín et al. (2015).
-

AUV photomosaics of the seafloor at the shallow-water Milos hydrothermal system. Photomosaics were acquired with the Sparus II (Iqua, Girona) AUV and constructed from vertically acquired still images. Images were acquired in July 2019 at the Paleochori and Spathi Bays. Photomosaics are provided as geotiffs (UTM Zone 35). This dataset includes both original photomosaics, that show distortions due to AUV navigation drift, and georeferenced photomosaics, that have been registered with the associated drone photomosaics at these same areas.
-
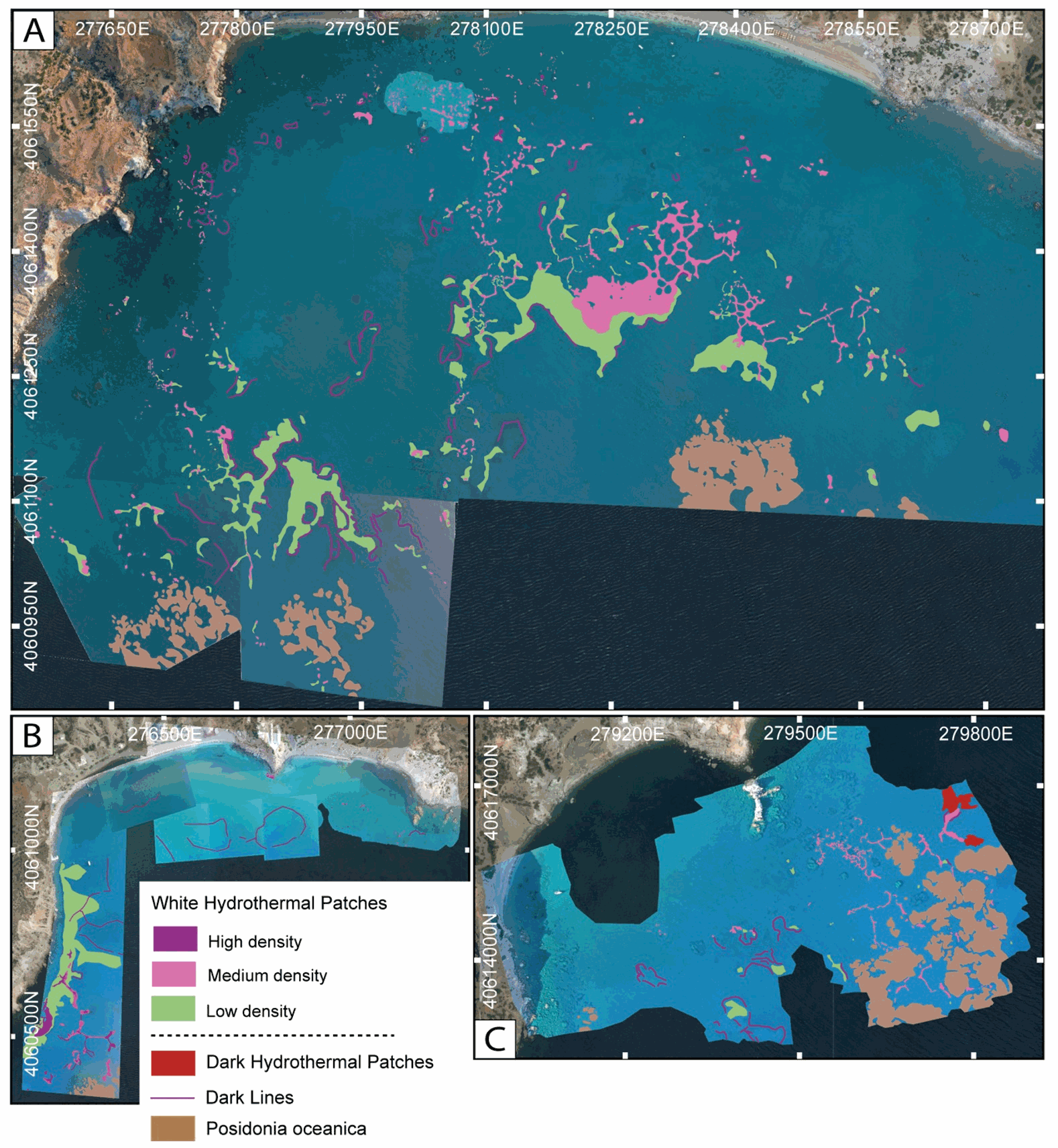
This dataset inscludes shapefiles interpreted from photomosaics over the Milos shallow water hydrothermal system, and instrument locaitons, from fieldwork in July and September 2019. The shapefiles are organized by study areas (Agia Kyria, Paleochori, and Spathi bays). Details are provided in the associated paper (Puzenat et al., 2021) in addition to information in the Readme File associated. a) Shapefiles of seafloor textures interpreted from drone imagery. b) Shapefiles of seafloor texttures interpreted from AUV imagery. c) Shapefiles of instruments deployed in the study area in September 2019.
-

Colour seafloor photomosaics along towed camera tracks crossing the axis of the Lucky Strike ridge segment (Mid Altantic Ridge). Images were acquired with the WHOI TowCam system (https://www.whoi.edu/what-we-do/explore/underwater-vehicles/towed-vehicles/towcam/), during the Graviluck 2016 cruise. Images are process for colour and illumination correction. Images are placed and photomosaiced based on USBL navigation of the TowCam, altitude, and orientation. The datasets include geotiffs for each of the tracks. To be cited: cited: Gini et al. (submitted); Escartin et al. (2008; 2014).
-
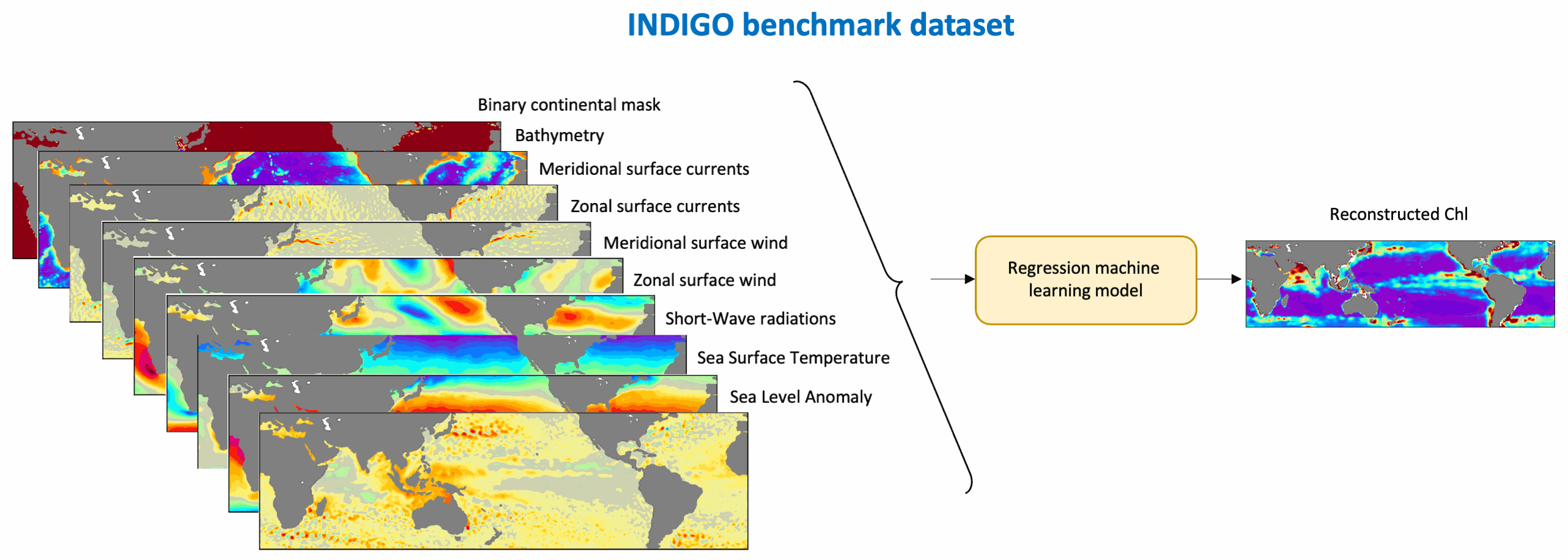
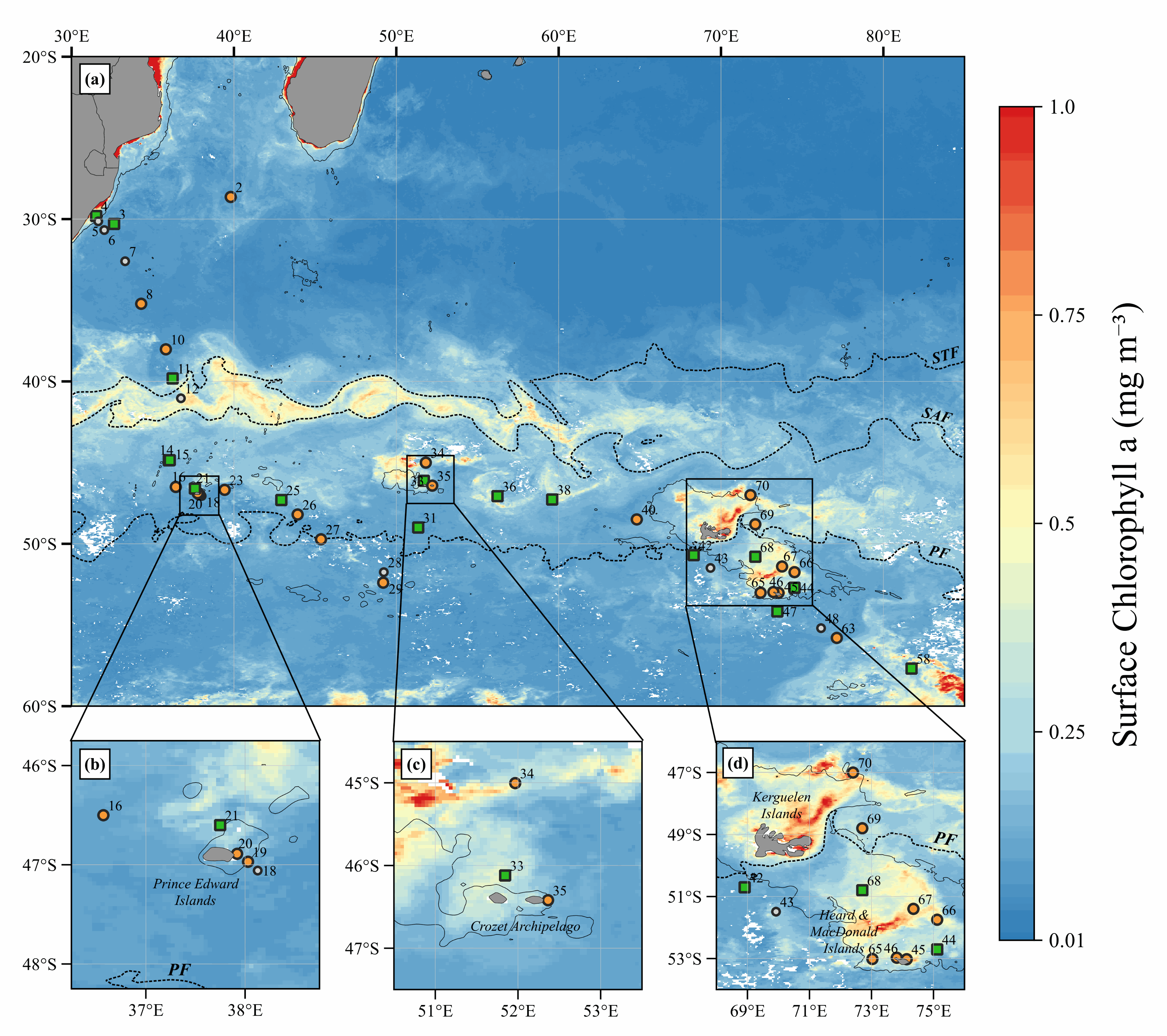
This benchmark dataset contains the physical data used as predictors to reconstruct global chlorophyll-a concentrations (Chl, a proxy of phytoplankton biomass) in Roussillon et al., as well as the reference satellite Chl target fields. The nine physical predictors' data (Short-Wave radiations, Sea Surface Temperature, Sea Level Anomaly, Zonal and meridional surface currents, Zonal and meridional surface wind stress, Bathymetry, Binary continental mask) were extracted from publicly available datasets over [1998-2015] and resampled to the same spatio-temporel resolution as Chl, i.e. monthly on a 1°x1° grid between 50°N and 50°S. Missing values were gap-filled using the heat diffusion equation. Each variable was normalized by substracting its mean from the original values and dividing by its standard deviation over [1998-2015]. This dataset was used to train and validate the Multi-Mode Convolutional Neural network (CNNMM8) introduced in Roussillon et al. ; reconstructed monthly Chl fields over the [2012-2015] test period are also provided here. We hope this benchmark dataset can help to promote the improvements of methods as well as the emergence of new ideas, as building datasets is sometimes more time-consuming than the implementation of machine learning tools themselves. This would also facilitate the quantitative comparison of models performances' on the exact same datasets.
-

The Capelinhos hydrothermal vent site is a large edifice of 588 m² from its base. Located off axis, 1.5 km east of the central lava lake of the main Lucky Strike Hydrothermal Vent Field (mLSHVF), it is characterised by distinct geochemical and thermal conditions from the others mLSHVF edifices, with notably high iron concentrations compared to sites around the lava lake. The edifice has been surveyed with the remotely operated vehicle Victor6000 for inter-annual monitoring during the 2023 Momarsat cruise. Video transects were collected during dive 844-6 with a forward-looking camera and video sequences were used to build 3D models of the edifice using structure-from-motion techniques. A first dataset includes 2971 images extracted every 2 seconds from the videos. The images are time-stamped and linked to a navigation file, which is generated by interpolating navigation data at the corresponding timestamps. A second dataset includes a 3D textured model of the site (obj format) together with the a kml file for georeferencing. The ROV navigation file (.txt) is provided separately. Point : 37.28948183 (lat) -32.26413676 (long) -1663.487 (depth)
-

The Sintra hydrothermal vent site reconstructed in this dataset is 311.82 m². It is located in the North east of the main Lucky Strike Hydrothermal Vent Field (LSHF). The edifice has been surveyed with the remotely operated vehicle Victor6000 for inter-annual monitoring during the 2023 Momarsat cruise. Video transects were collected during dive 844-6 with a forward-looking camera and video sequences were used to build a 3D model of the edifice using structure-from-motion techniques. A first dataset includes 1561 images extracted every 2 seconds from the videos. The images are time-stamped and linked to a navigation file, which is generated by interpolating navigation data at the corresponding timestamps. A second dataset includes a 3D textured model of the site (obj format) together with the a kml file for georeferencing. The ROV navigation file (.txt) is provided separately. Point : 37.2921244612 (lat) -32.2750499167 (long) -1612.34 (depth)