 Metadata catalogue
Metadata catalogue
2024
Type of resources
Available actions
Topics
INSPIRE themes
Keywords
Contact for the resource
Provided by
Years
Formats
Representation types
Update frequencies
status
Service types
Scale
Resolution
-
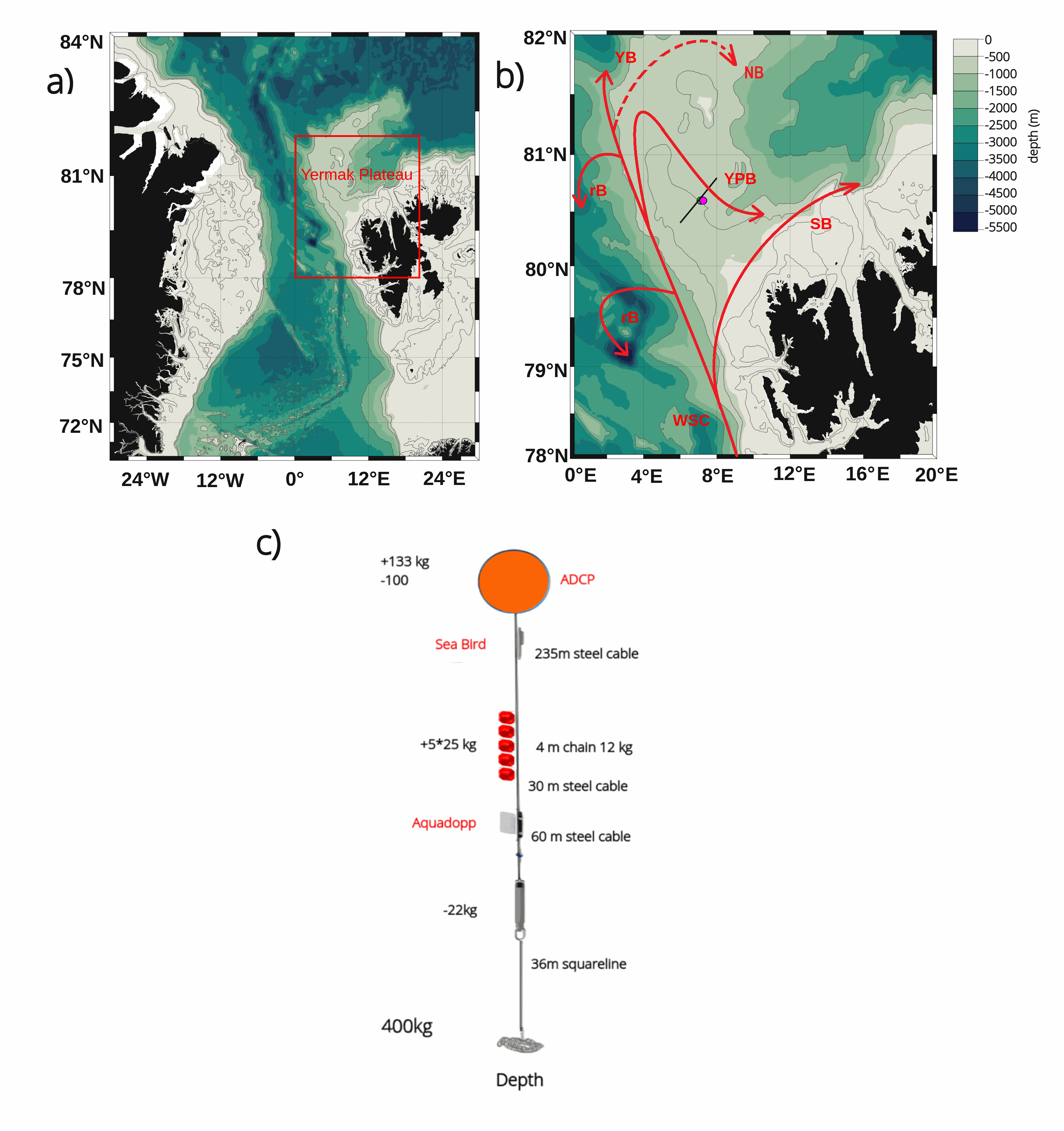
The mooring was deployed on 15 September 2017 from Norwegian Research Vessel Lance at 80.6°N and 7.26°E (depth of 730 m) in the Yermak Pass over the Yermak Plateau north of Svalbard. It comprised 3 instruments: an upward-looking RDI 75kHz, a Long Ranger Acoustic Doppler Current Profiler (ADCP) at 340 m with 16 m vertical resolution (25 bins of 16 m each) and a 2-hour sampling time; a Seabird SBE37 measuring temperature, salinity and pressure at 348 m with 10-minute sampling time; and an Aquadopp current meter at 645 m with a 2-hour sampling time. The mooring was retrieved on the 19 July 2020 by Norwegian Icebreaker K.V. Svalbard. The present dataset features: (i) the ADCP 50-hour smoothed daily velocities, conservative temperature and pressure time series interpolated every 10 meters within the 20-330m layer, (ii) the Aquadopp 50-hour smoothed daily velocities and pressure time series at 645 m; and (iii) the SBE37 50-hour smoothed daily conservative temperature, absolute salinity and pressure time series at 348 m.
-
Dataset from seabed mapping within the MAREANO programme, a collaboration between the Institute of Marine Research, the Geological Survey of Norway and the Norwegian Mapping Authority, Hydrographic Service. MAREANO maps depth and bathymetry, sediment composition, biodiversity, habitats and biotopes, and also pollution in the seabed within Norwegian offshore areas.
-
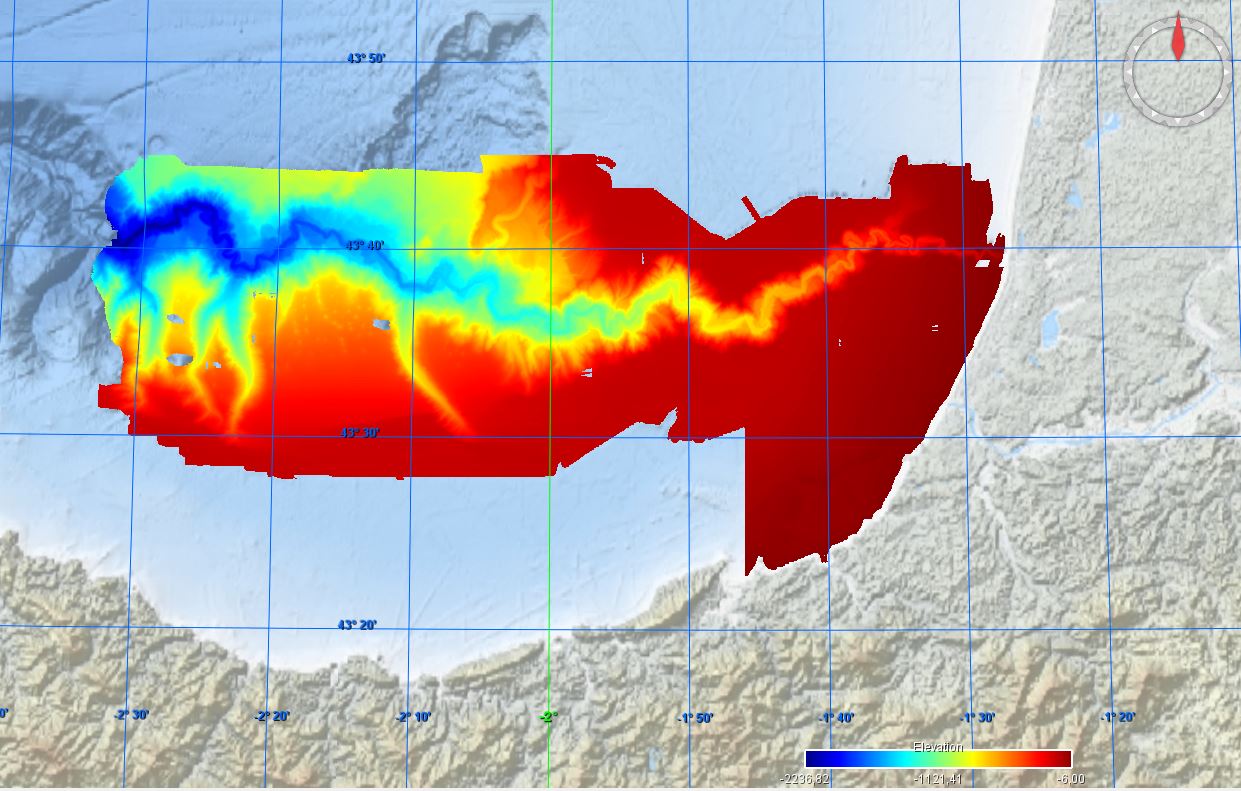
Conversion into the EMODnet format of the published grid for the Capbreton Canyon in 2007: http://dx.doi.org/10.12770/72e2f750-c255-11df-a9b6-005056987263
-
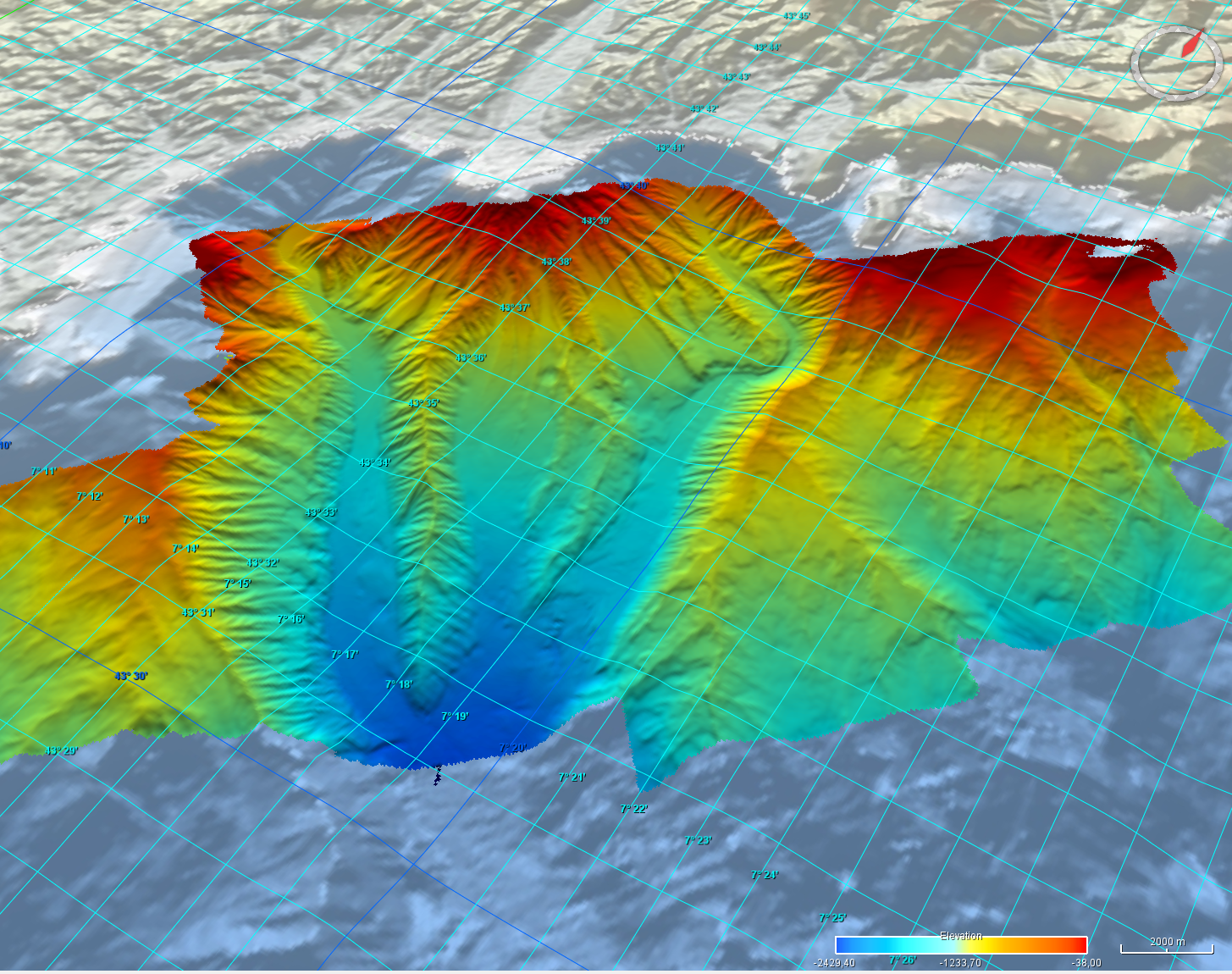
Grid processed for the purpose of the HR DTMs layer of EMODnet Bathymetry HRSM, October 24
-
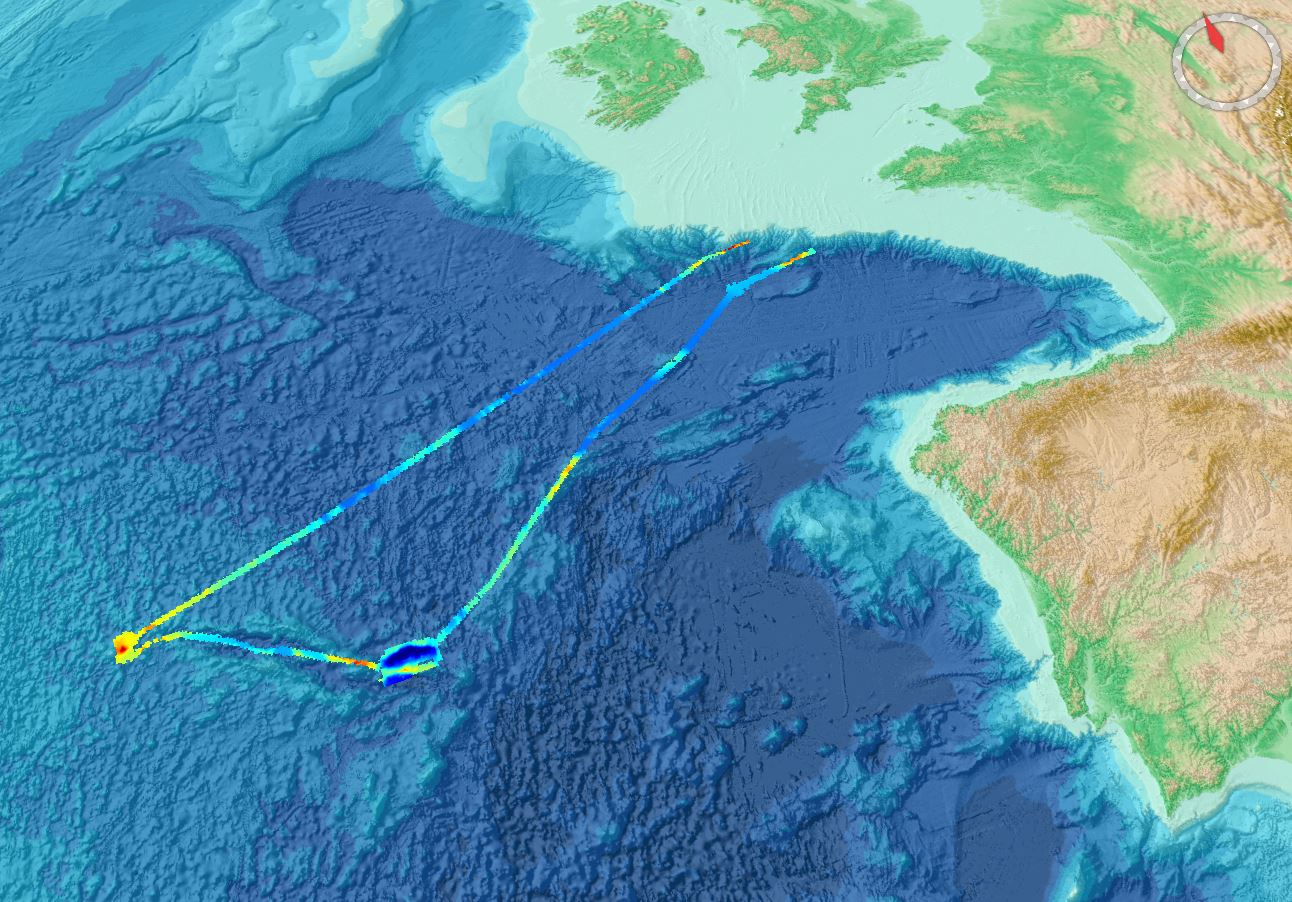
Gridded data for the ESS-ULYX-PP-2023-2 cruise.
-
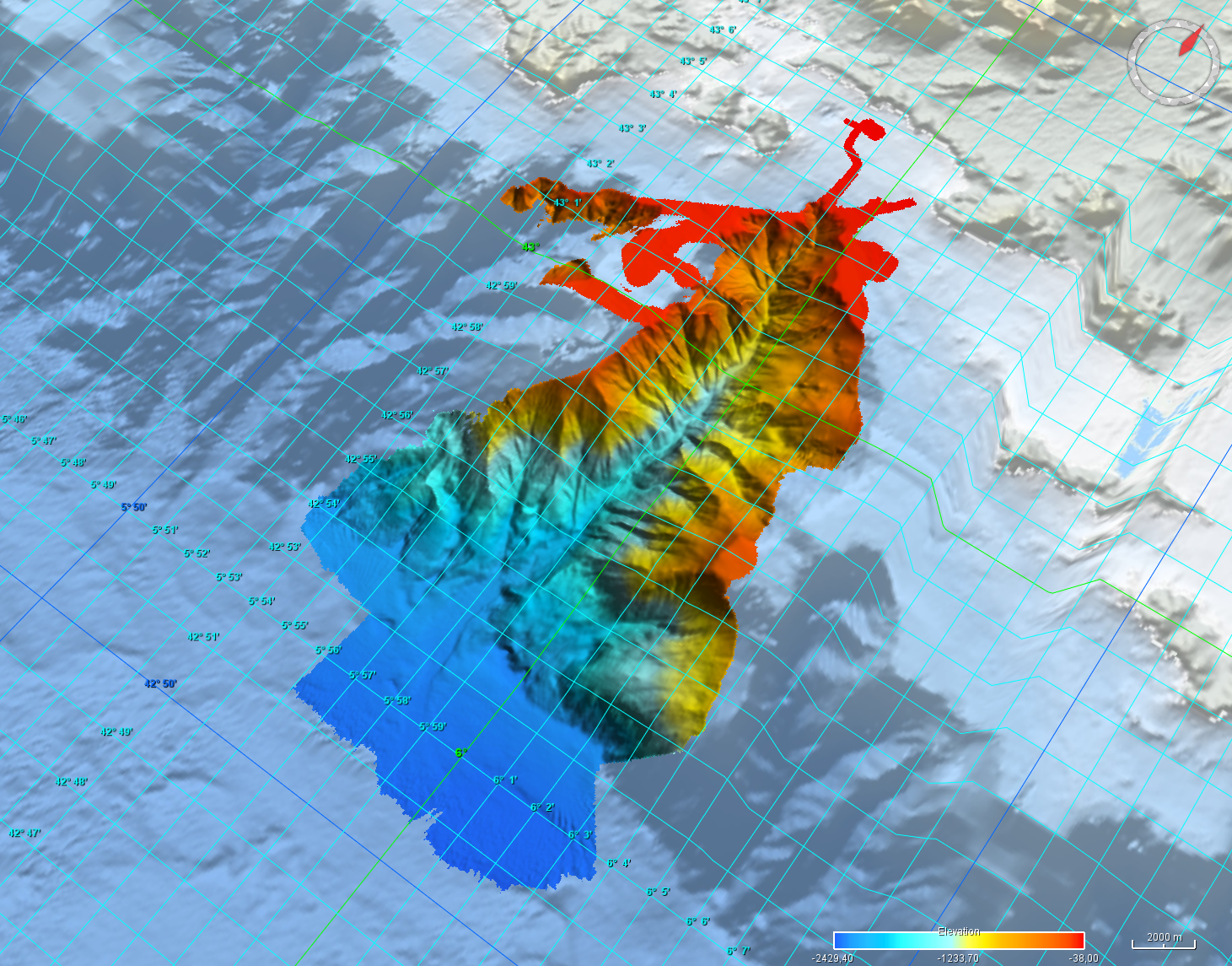
Grid processed for the purpose of the HR DTMs layer of EMODnet Bathymetry HRSM, October 2024
-
Sea fans (order : Alcyonacea) are the most remarkable and easily identifiable species of the subtidal hard substrate communities (Gili and Ros, 1985). From an ecological point of view, sea fans play an essential role (Gili and Coma, 1998) by increasing the biomass and the diversity of hard substrates (Mitchell et al, 1992; Ballesteros, 2006) via an umbrella species role providing habitat for small epifauna and a refuge for many fish (Ross and Quatrini, 2007). When their density is high enough, they form animal forests (sensu Rossi et al 2017) and become engineering species by modifying the level of turbulence and therefore of sedimentation of propagules in the benthic boundary layer (Chamberlain and Graus, 1975). In the shallow rocky habitats of the Mediterranean, five species of sea fans dominate: the white gorgonian Eunicella singularis (Esper, 1791), the orange gorgonian Leptogorgia sarmentosa (Esper, 1789), the yellow gorgonian Eunicella cavolinii (Koch, 1887), the red gorgonian Paramuricea clavata (Risso, 1826), and the red coral Corallium rubrum (Linnaeus, 1758). Some of these species, characterized by long life span, have been included in the management plans of the Gulf of Lion Marine Protected Areas, with surveys for the monitoring of the demographic structure of E. Singularis (Réserve Naturelle Marine de Cerbère-Banyuls, Aire Marine Protégée Agathoise) and C. rubrum (Réserve Naturelle Marine de Cerbère-Banyuls, Parc Marin de la Côte Bleue). In parallel with these management-related surveys, some scientific studies on the demographic structure and ecology of P. clavata, E. singularis and C. rubrum, with the aim of documenting mass mortality events (Cerrano et al. al., 2000, Garrabou et al., 2001) have been undertaken in the Parc Marin de la Côte Bleue, in the Parc National de Port-Cros, along the Côte Vermeille and Cap de Creus coast and in the Cinque Terre National Park (Garrabou and Harmelin, 2002, Torrents et al., 2005 ; Linares et al. 2008 Linares et al., 2010 ; Santangelo et al., 2011; Gori et al., 2011a ; Gori et al. 2011b; Rossi et al., 2008). The present database gathers two extensive inventory of sea fans populations performed between 2013 and 2015 in the Gulf of Lion and between 2018 and 2019 in the Ligurian Sea. The same protocol was applied to estimate the population density of the five species in 585 stations. The 585 stations were defined a priori on a regular mesh mapping the main hard-bottom substrate units of the Gulf of Lion (Côte Bleue, Plateau des Aresquiers, Plateau du Cap d'Agde, Cap Leucate, Côte Vermeille), and surrounding the ports of Toulon, La Spezia and Bastia (Figure 1). The spacing between stations was varied from 100m to 800 m according to bathymetry steepness. Each station was geo-referenced from the surface and located on the sea bed with a mooring. Counts of individuals of the five species were made by scuba divers trained to species identification in four quadrats (1m x 1m), positionned at 5 m from the mooring along the sea bed, in the four cardinal directions. In total, the sampling required 1500 dives. References Ballesteros E. 2006. Mediterranean coralligenous assemblages: a synthesis of present knowledge Oceanography and Marine Biology: An Annual Review 44, 123-195 Carpine C, Grasshoff M. 1975. Les gorgonaires de la Mediterranee. Bull Inst Oceanogr Monaco 71:1–140 Cerrano, C., Bavestrello G., Bianchi C.N., Cattaneo R. Vietti, S. Bava, C. Morganti, C. Morri, P. Picco, G. Sara S. Schiaparelli S. Siccardi A. & Sponga, F. 2000. A catastrophic mass-mortality episode of gorgonians and other organisms in the Ligurian Sea (Northwestern Mediterranean), summer 1999. Ecology Letters 3: 284–293. Chamberlain J.J. A., Graus, R.R. 1975. Water Flow and Hydromechanical Adaptations of Branched Reef Corals. : Bulletin of Marine Science 25 (1): 112-125 Garrabou J., Perez T., Sartoretto S., Harmelin J.G., 2001. Mass mortality event in red coral Corallium rubrum populations in the Provence region (France, NW Mediterranean). Mar Ecol Prog Ser 217:263–272 Garrabou J., Harmelin G. 2002. A 20-year study on life-history traits of a harvested long-lived temperate coral in the NW Mediterranean: insights into conservation and management needs. J Anim Ecol 71:966–978 Gili, J.M., & Ros, J. 1985. Study and cartography of the benthic communities of the Medes Islands (NE Spain). Marine Ecology 6, 219–238. Gili J.M. and Coma R. 1998. Benthic suspension feeders: their paramount role in littoral marine substrates. TREE 13 (8): 316-321 Gori, A., Rossi, S., Linares, C., Berganzo, E., Orejas, C., Dale, M. & Gili, J.M. 2011a. Size and spatial structure in deep versus shallow populations of the Mediterranean gorgonian Eunicella singularis (Cap de Creus, Northwestern Mediterranean Sea)” Marine Biology, DOI: 10.1007/s00227-011-1686-7 Gori A., Rossi S., Berganzo E., Pretus J.L., Dale M.R.T., Gili J.M. 2011b. Spatial distribution patterns of the gorgonians Eunicella singularis, Paramuricea clavata and Leptogorgia sarmentosa (Cape of Creus, Northwestern Mediterranean Sea). Mar Biol 158:143-158 Linares C, Coma R, Garrabou J, Diaz D, Zabala M. 2008. Size distribution, density and disturbance in two Mediterranean gorgonians: Paramuricea clavata and Eunicella singularis. J Appl Ecol 45:688–699 Linares C., Bianchimani O., Torrents O., Marschal C., Drap P., Garrabou J., 2010. Marine Protected Areas and the conservation of long-lived marine invertebrates : the Mediterranean red coral. Marine Ecology Progress Series 402: 69-79. Mitchell, N. D., Dardeau, M. R., Schroeder,W. W., Benke, A. C. 1992. Secondary production of gorgonian corals in the northern Gulf of Mexico. Mar Ecol. Prog. Ser. 87: 275-281 Ross S., Quattrini A. 2007. The fish fauna associated with deep coral banks off the southeastern United States. Deep Sea Research I (54): 975-1007. Rossi S, Bramanti L, Gori A, Orejas C. 2017. An Overview of the Animal Forests of the World. In book: Marine Animal Forests, pp.1-26. SPRINGER.[DOI: 10.1007/978-3-319-17001-5_1-1] Santangelo G., Bramanti L., Rossi S., Tsounis G., Vielmini I., Lott C., Gili J.M. 2011. Spatial patterns of variation in recruitment and post-recruitment processes of the Mediterranean precious gorgonian coral Corallium rubrum. Journal of Experimental Marine Biology and Ecology. DOI: 10.1016/j.jembe.2011.10.030 Torrents O., Garrabou J., Marshal C., Harmelin J.G., 2005. Age and size at first reproduction in the commercially exploited red coral Corallium rubrum (L.) in the Marseilles area (France, NW Mediterranean). Biol Conserv 121: 391–397 Rossi S., Tsounis G., Orejas C., Padrón T., Gili J.M., Bramanti L., Teixidó N. and Gutt J. 2008. Survey of deep-dwelling red coral (Corallium rubrum ) populations at Cap de Creus (NW Mediterranean). Marine Biology 154 (3): 533-545.
-

This seismic data set was recorded in September 2015 on board the R/V Pourquoi Pas? during the GHASS cruise (IFREMER) on the western Black Sea, offshore Romania. The available profiles (part of the collected data) within this deposit are located on the continental slope, bathymetry between 500 to 1200 meters. The pre-processing of the seismic data included common midpoint binning @6.25 meters, trace and shot edition, source delay correction, and a 35-375 Hz band pass filtering. Detailed Root Mean Square Velocity analyses were performed on semblance panels computed using super gathers every 150 m. Normal Move Out time correction was then applied on the Common Mid Point (CMP) using these velocities prior to stack. Interval velocities were computed using the Dix equation. The velocity model was then interpolated every CMP location, converted to depth and smoothed to perform post-stack depth migration. The depth migrated sections and the depth velocity models have been output to standard SEG-Y rev1 format (https://library.seg.org/pb-assets/technical-standards/seg_y_rev1-1686080991247.pdf) with values written using “big-endian” byte ordering and IEEE floating-point. For a given profile, both SEGY files have the same number of traces and the same bin locations. Velocity unit is in meter.second-1. The depth sampling is set to 0.5 meter for both files. The recording delay is zero for the depth migration SEGY files. The delay is coded in meters and constant for a given depth velocity SEGY file, stored within the Trace Header (bytes 109-110). Trace coordinates are also stored within the Trace Header using WGS84 +DDDMMSS.ss format with a scale factor of -100 (bytes 81-88, which means that the value has to divided by 100) . For more convenient access to the location of the profiles, these coordinates are also stored into ASCII files using decimal degrees.
-

The coastal topo-bathymetric product of a part of the Norman-Breton Gulf at a resolution of 0.0002° (~ 20 m) was produced within the framework of the PAPI Saint-Malo (with the financial assistance of Saint-Malo Agglomération, the council department of Ille-et-Vilaine, of the Ministry of Ecological Transition and Shom). This coastal DEM encompasses the Minquiers plateau in the northwest to the bay of Mont-Saint-Michel in the southeast. This DTM is intended to be implemented in hydrodynamic models in order to produce forecasts as precise as possible of water levels and sea states at the coast and therefore to improve the management of the risk of marine submersion. This DTM is offered openly on the Shom diffusion website. https://dx.doi.org/10.17183/MNT_COTIER_GNB_PAPI_SM_20m_WGS84
-
Arctic Western Eurasian Basin: IAOOS 23 and IAOOS 24 ocean CTD-DO, CDOM and nitrate profiles in 2017
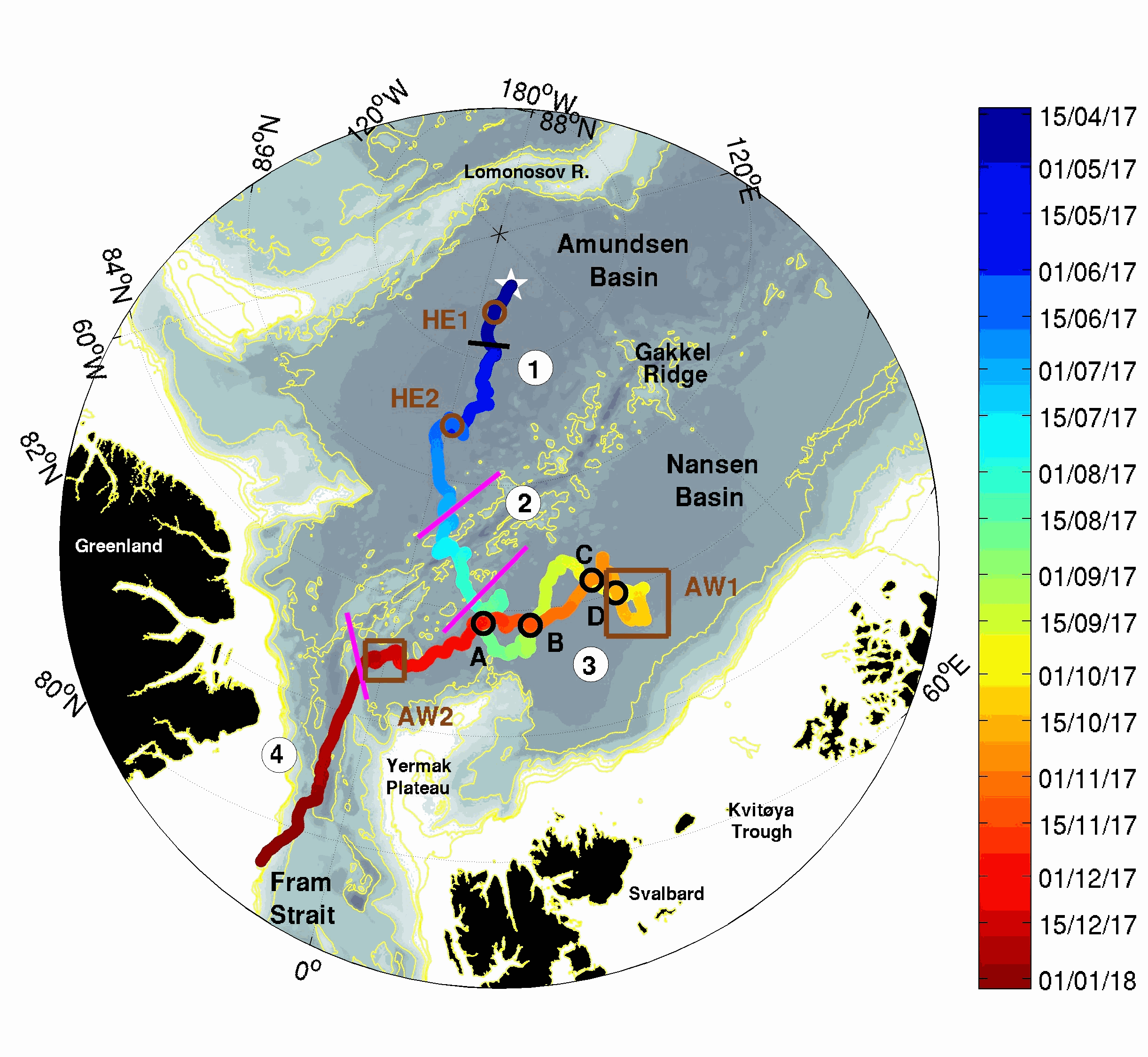
The two platforms IAOOS 23 and IAOOS 24 were deployed within 600 m from each other at the North Pole from the Russia-operated Barneo ice camp on April 12, 2017. They followed a meandering trajectory, reaching as far as 30°E in the Nansen Basin, before turning back to the western Fram Strait. On both IAOOS 23 and 24, the ocean profiler was a PROVOR SPI (from French manufacturer NKE) equipped with a Seabird SBE41 CTD (Conductivity, Temperature, Depth) and a dissolved oxygen (DO) Aandera 4330 optode. For the first time, the profiler on IAOOS 23 also carried biogeochemical sensors. It featured a bio-optics sensor suite and a submersible ultraviolet nitrate analyzer (SUNA, Satlantic-Seabird Inc.). The bio-optics sensor suite (called Pack Rem A) combines a three-optical-sensor instrument (ECO Triplet, WET Labs Inc.) and a multispectral radiometer (OCR-504, Satlantic Inc.). The present dataset is composed of CTD-DO data from IAOOS 23 and 24, corrected from the thermal lag and the sensor lag, despiked and interpolated vertically every 0.5 m. It also comprises nitrate concentrations from the SUNA and CDOM fluorescence from the WETLabs ECO sensor on IAOOS 23. Other biogeochemical data will be added to this dataset. The profilers were set to perform two upward profiles a day from 250 m (IAOOS 23) and 350 m (IAOOS 24) upward starting at approximately 6 am and 6 pm. They provided a unique 8-month long dataset, gathering a total of 793 profiles of the temperature, salinity and oxygen (upper 350m) and 427 profiles of CDOM and nitrates concentrations (upper 250m).